Comparative Environmental Physiology (CEPh)
Research Interests
Climate change and biogeography
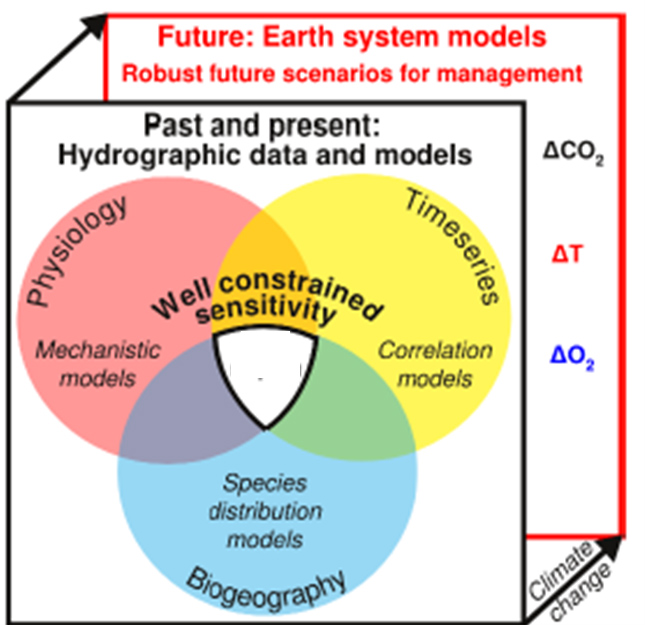
Climate and physiology shape biogeography, yet the range limits of species have rarely been ascribed to the quantitative traits of organisms. We compare metabolic metrics to geographical range boundaries for a global cross-section of the biodiversity of marine animals. For all studied species, thermal and hypoxic limits are substantially reduced by the energetic demands of ecological activity. The ratio of active to resting oxygen demand found at the geographic range boundaries is similar across species. Thus, active temperature-dependent oxygen supply links the biogeography of diverse marine species to fundamental energetic requirements that are shared across the animal kingdom, providing a benchmark that can be used to predict shifts in species distributions with climate change.
Select publications:
Deutsch, C., J. L. Penn, and B. A. Seibel. 2020. Metabolic trait diversity shapes marine biogeography. Nature. DOI: 10.1038/s41586-020-2721-y.
Howard, E. H., J. L. Penn, H. Frenzel, B. A. Seibel, D. Bianchi, L. Renault, F. Kessouri, M. A Sutula, J. C. McWilliams, and C. Deutsch. 2020. Climate driven aerobic habitat loss in the California Current System. Science Advances. 6: eaay3188.
Deutsch, C., Ferrel, A., Seibel, B. A., H. O. Pörtner, R. B. Huey. 2015. Climate change tightens a metabolic constraint on marine habitats. Science 348: 6239 pp. 1132-1135. DOI: 10.1126/science.aaa1605
Oxygen supply and demand
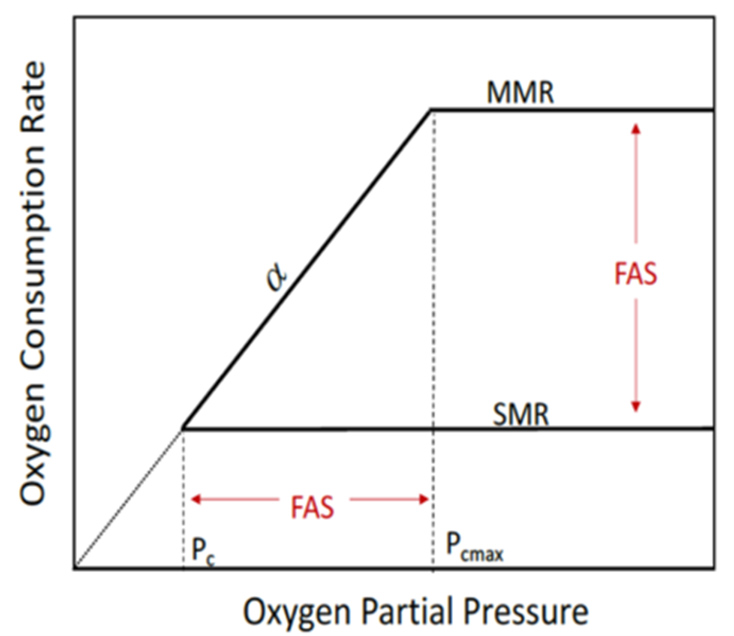
The capacity to extract oxygen from the environment and transport it to respiring tissues in support of metabolic demand may have implications for species’ thermal tolerance, body size, diversity and biogeography. A recently discovered linkage between maximum and basal metabolic rate and their oxygen, temperature and size dependencies suggests that the physiological capacity for oxygen supply (α) precisely matches the maximum evolved demand at the highest persistently available oxygen pressure. For most terrestrial and shallow-living marine species, this is the current atmospheric pressure, 21 kPa. Any reduction in oxygen partial pressure from current values will result in a quantifiable decrement in aerobic performance. This new relationship redefines many important physiological concepts and alters their ecological interpretation.
Select publications:
Seibel, B. A. and C. Deutsch. 2020. Oxygen supply capacity in animals evolves to meet maximum demand at the current oxygen partial pressure regardless of size or temperature. J. Exp. Biol. 10.1242/jeb.210492
Seibel, B. A., Andres, A., Birk, M. A., Shaw, C. T., Timpe, A. and C. Welsh. 2021. Oxygen supply capacity breathes new life into the critical oxygen partial pressure (Pcrit). Journal Experimental Biology.
Seibel, B. A., Häfker, S., Trübenbach, K., Zhang, J., Pörtner, H. O., Rosa, R. and Storey, K. B. 2014. Metabolic suppression during protracted exposure to hypoxia in the jumbo squid, Dosidicus gigas, living in an oxygen minimum zone. J. Exp. Biol. 217, 2555-2568
Ocean Deoxygenation
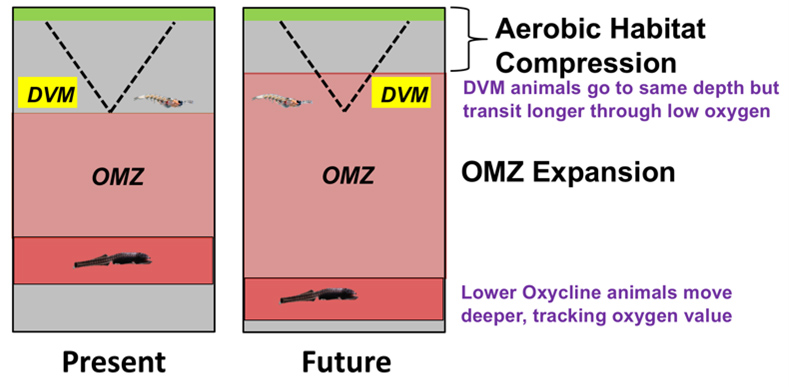
Animals are directly dependent on the availability of oxygen for aerobic metabolism. Diversity, abundance, distribution and composition of species are all influenced by variations in oxygen at both large and small scales. Ocean deoxygenation will decrease the minimum oxygen content in the mesopelagic zone and cause oxyclines to shift vertically (i.e. expansion of the OMZ core) in the water column. Changes in temperature and oxygen profiles within the water column may therefore alter species composition, ecology and diversity. These changes may substantially alter the biogeochemical cycles (e.g. the biological pump) that rely on the mesopelagic community.
Select publications:
Wishner, K. F., B. A. Seibel, C. Roman, C. Deutsch, D. Outram, C. T. Shaw, M. A. Birk, K. A. S. Mislan, T. J. Adams, D. Moore, S. Riley. 2018. Ocean deoxygenation and zooplankton: Very small oxygen differences matter. Science Advances 4, eaau5180. Paper of the Year, Deep-sea Biology Society.
Seibel, B. A. and Wishner, K. F. 2019. Ch. 8.1 Mesopelagic Community response to deoxygenation. In IUCN: Ocean deoxygenation: Everyone’s problem. Laffoley, D. and J. M. Baxter, eds.
Seibel, B. A., Häfker, S., Trübenbach, K., Zhang, J., Pörtner, H. O., Rosa, R. and Storey, K. B. 2014. Metabolic suppression during protracted exposure to hypoxia in the jumbo squid, Dosidicus gigas, living in an oxygen minimum zone. J. Exp. Biol. 217, 2555-2568
Ocean Acidification
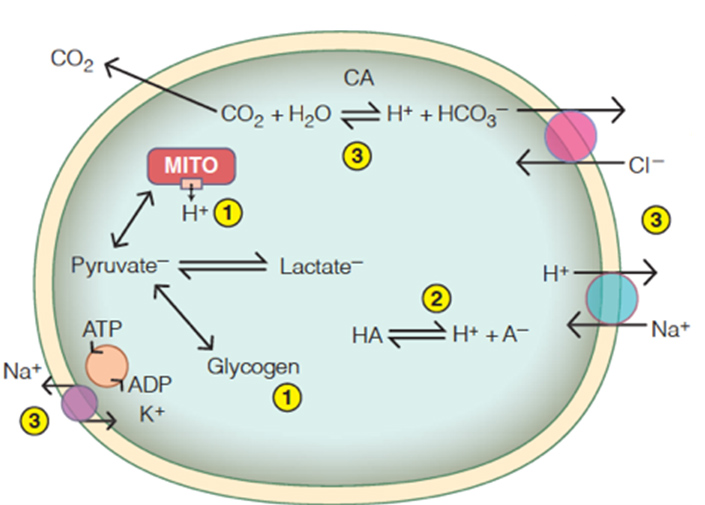
Atmospheric carbon dioxide partial pressure (PCO2) has increased from the pre-industrial mean of 280 ppmv to over 400 μatm today and may reach 1000 μatm by the year 2100. Anthropogenic CO2 also diffuses into the ocean, where it reacts with water, resulting in reduced pH. This phenomenon, known as ocean acidification (OA), may pose a serious problem for marine organisms. Those with calcareous skeletons or shells, such as corals and some plankton, have been at the center of scientific interest. However, OA may also have detrimental effects on the survival, growth, and physiology of marine animals more generally. We study such effects in diverse species living across regions and ecosystems.
Select publications:
Melzner, F., L. Tomanek and B. A. Seibel. 2020. Ocean acidification impacts on coastal invertebrates. Frontiers in Marine Science. Ann. Rev. Mar. Sci. 12:12.1–12.25
Birk, M. A., McLean, E. L. and B. A. Seibel. 2018. Ocean acidification does not limit squid metabolism via blood oxygen supply. J. Exp. Biol. 221, jeb187443. doi:10.1242/jeb.187443.
Rosa, R., and Seibel, B. A. 2008. Synergistic effect of climate-related variables suggests future physiological impairment in a top oceanic predator. Proc. Nat. Acad. Sci. 52. 20776-20780.
Seibel, B. A. and P. J. Walsh. 2001. Potential impacts of CO2 injection on deep-sea biota. Science. 294(5541): 319-320.
Temperature Adaptation
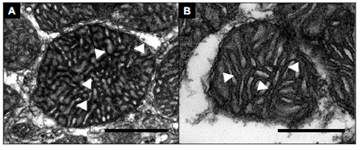
In ectotherms living in cold waters, rates of energy consumption are comparatively slow. At -1.8 °C, the waters of Antarctica pose a formidable physiological barrier for most ectotherms. Conversely, warm waters pose a potential constraint from, among other limitations, reduced oxygen supply relative to demand. We study the specific neuromuscular and metabolic adjustments that enhance metabolic capacity in marine organisms in environments from the equator to the poles. We’ve shown that polar animals have undergone substantial tissue-level reorganizations to accommodate their environment, which may reduce their capacity to acclimate to a changing climate. Conversely, oxygen supply capacity increases with temperature to match high demands in tropical waters.
Select publications:
Slesinger, E., A. Andres, R. Young, B. A. Seibel, V. Saba, B. Phelan, J. Rosendale, D. Wieczorek, G. Saba. 2019. The effect of ocean warming on black sea bass (Centropristis striata) aerobic scope and hypoxia tolerance. PLoS ONE 14(6): e0218390
Dymowska, A., Manfreddi, T., Rosenthal, J. C., and B. A. Seibel. 2012. Muscle ultrastructure and mitochondrial morphometrics in polar and temperate pteropods (Gymnosomata: Gastropoda). J. Exp. Biol. 215: 3370-3378.
Rosenthal, J. C., Seibel, B.A., Dymowska, A., and F. Bezanilla. 2009. Trade-off between aerobic capacity and locomotory activity in an Antarctic pteropod. Proc. Natl. Acad. Sci. 106: 6192-6196.
Pelagic Ecology and Metabolic Variation

The rates of metabolism in animals vary tremendously throughout the biosphere. The origin of this variation is a matter of active debate: some scientists highlight the importance of anatomical or environmental constraints while others emphasize the diversity of ecological roles and the associated energy demands. Across a depth range, metabolic rate varies by more than 200-fold, independent of body mass and temperature. This suggests that high metabolic demand follows strong selection for locomotory capacity among visual predators inhabiting well-lit oceanic waters. Relaxation of such selection in the deep-sea, where visual predation is limited by lack of light, provides an opportunity for reduced energy expenditure. Large-scale metabolic variation in the ocean results from interspecific differences in ecological energy demand.
Select publications:
Seibel, B. A. and Drazen, J. C. 2007. The rates of metabolism in marine animals: environmental constraints, ecological demands and energetic opportunities. Phil. Trans. Roy. Soc. Lond. B. 362: 2061-2078.
Seibel, B. A. 2007. On the depth and scale of metabolic rate variation: scaling of oxygen consumption and enzymatic activity in the Class Cephalopoda. J. Exp. Biol. 210: 1-11.
Seibel, B. A. 2016. Cephalopod susceptibility to asphyxiation via ocean incalescence, deoxygenation and acidification. Physiology. 31: 418–429.
Life-history evolution

Deep-sea and open-ocean habitats are some of the least explored on Earth. We are interested in the animals that inhabit these regions and their physiological and behavioral adaptations to extreme environments. Interest in industrial activities such as deep-sea mining highlights the importance of understanding biological processes in these regions of the ocean. For example, reproduction is key to survival in any environment and yet the reproductive biology of most open-ocean and deep-sea species is unknown. Through our observations, we’ve described a novel egg-brooding behavior in deep-sea squids, giant floating egg “spheres” from the jumbo squid, and observed the longest egg-development period of any animal (more than four years) in a deep-sea octopus. These discoveries provide a glimpse into the life histories of animals in these remote habits.
Select publications:
Seibel, B. A., Robison, B. H., Haddock, S. D. H. 2005. Post-spawning egg-care by a squid. Nature 438: 929.
Robison, B. H., Drazen, J. C., and Seibel, B. A. 2014. Protracted egg-brooding in a deep-sea octopus. PLoS ONE. 9(7): e103437.
Birk, M. A., Paight, C. and Seibel, B. A. 2017. Observations of multiple pelagic egg masses from small-sized jumbo squid (Dosidicus gigas) in the Gulf of California. Journal of Natural History. 51: 2569-2584. DOI: 10.1080/00222933.2016.1209248
Trimethylamine Oxide
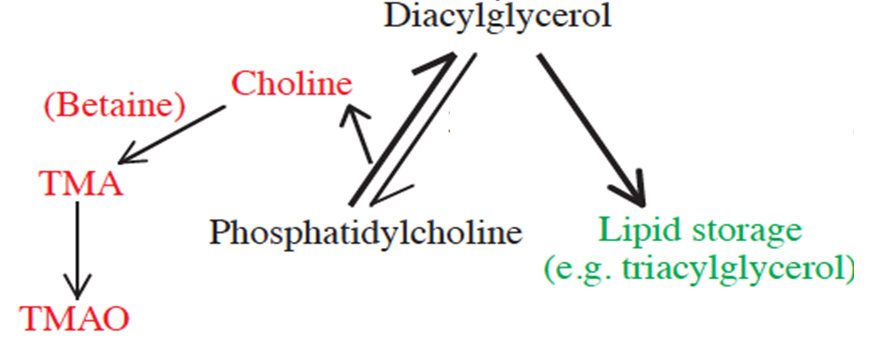
Trimethylamine oxide (TMAO) is a common and compatible osmolyte in muscle tissues of marine organisms. It is often credited with counteracting protein-destabilizing forces, such as hydrostatic pressure, in deep-sea organisms. Recent interest in TMAO stems from its role in cardiovascular disease and other pathologies. However, the origin and synthetic pathways of TMAO remain unclear. Marine animals are viable models for study of TMAO dynamics due high intrinsic TMAO levels. The hydrolysis of the common membrane lipid phosphatidylcholine, results in diacylglycerol which is stored as fat, and choline, which is converted to TMAO. Thus, TMAO is highest in animals that store large amounts of fat, such as deep-sea species. Whether TMAO is a specific adaptation to counteract pressure and other cellular stressors, or is merely a beneficial side product of lipid synthesis, remains unclear.
Select publications:
Bockus, A. and B. A. Seibel. 2016. Trimethylamine oxide and lipid levels increase with depth in Hawaiian midwater fishes. Deep-sea Res 112: 37-44.
Seibel, B. A. and P. J. Walsh. 2002. Trimethylamine oxide accumulation in marine animals: relation to acylglycerol storage? J. Exp. Biol. 205(3): 297-306.
Samerotte, S. L., J. C. Drazen, G. L. Brand, B. A. Seibel, and P. H. Yancey. 2007. Correlation of trimethylamine oxide and habitat depth within and among species of teleost fishes: an analysis of causation. Physiol. Biochem. Zool. 80(2): 197-208.